Coat color, temperament and domestication of Norway rats and deermice
For an introduction to the topic of coat color, temperament and domestication, see the more general coat color, temperament, and domestication page. This page is intended as a more in-depth look at the experimental evidence in rats and deermice.
- The agouti gene
- Black rats are more docile than agouti rats: two studies
- The Keeler studies: Keeler 1942, Keeler and King 1947
- Cottle and Price 1981
- Black deermice are more docile than agouti deermice
- How the agouti gene regulates coat color
- How the agouti gene may influence behavior
- The agouti allele and rat domestication
- Black rats are more docile than agouti rats: two studies
- The hooded gene
Studies have identified two genes that influence rat temperament and may have played a role in rat
domestication: the agouti gene and the hooded gene.
The agouti gene in rats has two main variants (called alleles): the agouti allele and the nonagouti allele. Rats with the agouti allele (A) have hairs that are banded with yellow-red and black pigment. Rats with the nonagouti allele (a) have hairs that are solid black.
The agouti allele is associated with more than just coat color, however. Evidence suggests that the agouti allele may also affect behavior: in behavioral tests comparing agouti rats and nonagouti rats of similar genetic background, nonagouti rats are found to be more docile than agouti rats.
Black rats are more docile than agouti rats: three studies
The Keeler studies
Keeler (1942): agouti and black rats
The rats: Keeler (1942) obtained agouti and black rats by first crossing albino laboratory rats (homozygous for hooded and black) with wild agouti rats. The hybrid offspring were all agouti or agouti berkshires. These hybrid offspring were then crossed together, producing rats that were either agouti, black or albino, with solid, berkshire or hooded patterns. These rats were not handled after weaning, and were not allowed to become familiar with experimenters.
The behavioral tests: Keeler put these second-generation agouti (agouti plus agouti hooded) and black rats (black plus black hooded) through a variety of behavioral tests to study how docile they were.
The results: The results were striking: when the rats were picked up and gently held for the first time, the agouti rats were significantly more likely to display untame behaviors than the black rats: more agouti rats bit (40% of agouti rats vs. 10% of black rats), squealed (68% vs. 37%), laid their ears back (45% vs. 10%) and urinated (51% vs. 7%) than black rats did. In addition, only 1% of agouti rats showed no reaction upon being picked up, compared to 30% of black rats (Table 1).
More agouti rats than black rats responded to mild annoyance. When their noses were tickled lightly with a fine bristle brush, 86% percent of agouti rats bit, compared to only 20% of black rats. Agouti rats were also much more reactive: only 15% of agouti rats showed no reaction to the brush, compared to 73% of black rats (Table 2).
The intensity of the responses varied between the two colors as well. The squeals of the agouti rats were often loud and prolonged, while those of black rats were short and soft and tended to decline into a protesting whine. The urination of the agouti rats was a strong spurt or stream; the urination of the black rats was a slow emission of one or a few drops. Agoutis defecated three to five boluses of feces; black rats frequently defecated only a single bolus. Lastly, agouti biting entailed a series of six or seven forceful bites delivered with "machine-gun rapidity," while in the black rat the bite was often a single, weak nip.
The response to handling wasn't the only difference. Agouti rats were more cautious than black rats in emergence tests. When the cage door was opened, agouti rats took an average of 84 seconds to come out, while black rats took only 57 seconds. After three minutes, 49% of the agouti rats had emerged, compared to 73% of black rats.
On the whole, Keeler (1942) found significant differences in behavior between agouti and black rats, but no clear, sharp division between the animals of different coat color. This is to be expected, because temperament is not a single-gene trait but is influenced by many genes.
Keeler and King 1947: agouti and black rats
In a more observational study, Keeler and King (1947) describe a study in which wild agouti Norway rats were raised in captivity for 35 generations. Many mutations appeared spontaneously in this population over the years: albino, hooded, red-eyed dilute, black, curly, albino-waltzer, cinnamon, and stub.
Some of these mutations were associated with varying degrees of tameness. Keeler and King note that the black rats in particular were much tamer than the agoutis. The black rats are described as "Already tame by nature. If very excited they may click their teeth, but are not apt to bite." Keeler and King conclude that their strain of tame albino rat, which was homozygous for black and hooded, "was not domesticated by painstaking selection over long periods of time, but was modified in morphology principally by the introduction of three coat color genes [albino, hooded, and black], and in behavior principally by the (non-agouti) black gene."
Cottle and Price (1987)
The rats: Over forty years later, Cottle and Price (1987) conducted a more complex experiment comparing the behavior of agouti and black rats. These rats were the descendants of six wild rats trapped in New York. The original wild rats had been found to be heterozygous for agouti (Aa). Five generations later, the authors took 12 female and 10 male rats who were heterozygous for agouti (Aa), and bred them to produce agouti (A-) and black rats (aa). These sixth-generation rats therefore had a common genetic background, and were put through behavioral tests.
The behavioral tests and results: In handling tests, the agouti rats were more likely to exhibit defensive behavior and were more difficult to handle than black rats. Specifically, when an experimenter placed a gloved hand on the floor near the rat, a higher percentage of agouti than black rats attacked the hand, and exhibited jumping and running behavior. When the experimenter slowly approached a gloved hand and tried to touch, stroke, then catch the rat, agouti rats were more likely than black rats to attack the hand and were significantly more difficult to touch, stroke and catch (Table 3).
All black rats could be touched by the second trial, but after five trials 15% of agouti rats had not been touched. All black rats could be stroked by the third trial, but after five trials 32% of agouti rats had not been stroked. The first time a rat was approached with the gloved hand, 76% of black rats could be caught, compared to only 25% of the agoutis. By the fifth trial, all black rats could be caught, compared to 75% of agoutis (Table 3).
Cottle and Price also found agouti rats to be more active in an open-field test, and groomed slightly less than nonagouti rats when placed on a platform.
Black deermice are more docile than agouti deermice
Hayssen (1997) found behavioral differences between agouti and black deermice that are similar to those found in rats.
The deermice: Wild deermice were captured and brought into the laboratory in the 1970s. Some of these wild deermice were heterozygous for nonagouti. Homozygous agouti and nonagouti lines were established and were bred in the laboratory for over 20 generations under the same conditions. Hayssen (1997) conducted behavioral tests on the agouti and nonagouti deermice following the same methodology as Cottle and Price (1987).
The behavioral tests and results: In handling tests, more agouti deermice than black deermice attacked and bit the experimenter's hand. Agouti deermice also jumped more frequently than black deermice. Sixty-one percent to 75% of nonagouti deermice were catchable, compared to only 41-59% of agoutis. Nonagoutis were also consistently more touchable and strokeable than agouti deermice.
Agouti deermice defecated less and walked around more than nonagouti deermice in an open-field test. When placed on a platform, agouti deermice usually jumped off and started exploring, while nonagoutis stayed on the platform and groomed.
Therefore, nonagouti deermice were less aggressive, less active, and easier to handle than their agouti counterparts.
How the agouti allele regulates coat color
Agouti hairs are banded with two types of melanin: a light reddish-yellow pigment and a dark brownish black pigment. The relative proportion of these two types of bands on the hairs is controlled by a hormone called MSH (melanocyte-stimulating hormone).
These pigments are produced in melanocytes, which are found at the base of each hair. When left alone, melanocytes normally produce a reddish-yellow pigment. When MSH comes along, however, it stimulates the pigment cells to produce the brownish-black pigment, creating a black band on the hair.
The agouti protein is a molecule that stops MSH from stimulating the pigment-producing cells (Lu et al. 1994, Willard et al. 1995). So when the agouti protein is present it binds MSH, so MSH can't stimulate the pigment-producing cells. The unstimulated pigment cells produce the yellow-red pigment. When the agouti protein is absent, MSH has free rein and it stimulates the pigment cells to produce dark pigment. In agouti-colored rats, the interplay between the agouti protein and MSH produces hairs with yellow and black bands.
If there's a mutation in the agouti gene, it codes for a dysfunctional agouti protein that cannot bind MSH, leaving MSH totally free to bind to and stimulate the pigment cells. These pigment cells therefore produce continuous dark pigment. Therefore, rats with the nonagouti allele do not produce yellow pigment and are entirely black.
How the agouti allele may influence behavior
The agouti protein does more than regulate pigment production in hair. MSH and its family of melanocortins are found elsewhere in the body, including the brain, where they have been found to be potent neuromodulators with diverse effects on mammalian behavior and physiology (De Weid and Jolles 1982, O'Donohue and Dorsa 1982, Voisey et al. 2003). The behavioral effects of the agouti protein may therefore be mediated through melanocortin receptors on neurons, but exactly how this occurs is not yet understood.
In addition, agouti and nonagouti animals have strikingly different neural profiles of catecholamines. Catecholamines (like dopamine and noradrenaline) are neurotransmitters that activate the body and prepare it to deal with stress. The agouti protein affects the distribution of catecholamines in the brain, and hence may influence the animal's docility (Hayssen et al. 1994).
Lastly, nonagouti rats were found to have smaller adrenal glands than agouti rats, when a black mutant was crossed with a wild agouti rat and the offspring were bred together to produce a litter of agouti and black rats (Keeler et al. 1947). The adrenal glands are responsible for producing adrenaline, a stress hormone.
The agouti allele and rat domestication
Most behaviors are influenced by many genes, but the agouti gene appears to have a disproportionate impact on the behavior, and may therefore have played a role in the domestication of the rat.
The temperament difference between the original wild-type agouti and non-agouti rats may be one of the historic reasons that the non-agouti allele is so common today in domestic rats. Eighty percent of fifty domestic inbred laboratory rat strains are homozygous for the nonagouti allele, as are seventy percent of 140 domestic inbred laboratory mouse strains (Festing 1979a and b, Festing 1989, Staats 1981).
Non-agouti rats may have been easier to capture, and once captured they may have been easier to handle, and therefore preferred. This preference for calmer temperament would be in addition to any natural human preference for an unusually colored rat. In addition to biased capture and breeding, the early non-agouti rats may have been less stressed by captivity, so they survived and reproduced better than their agouti counterparts, regardless of human wishes.
The hooded gene
Piebaldness -- patches of depigmented, white areas on the coat -- is also connected to calm temperament. The most famous example of this relationship is found in Belyaev's foxes, which were bred for tameness for generations. This selection for tame behavior resulted in tame, piebald foxes (Belyaev 1979, Trut 1999).
In rats, the main genetic locus of piebaldness is the hooded locus, which has two main variants (alleles): the dominant wild type allele (H) produces a solid-colored animal. The recessive hooded allele (h) produces patches of depitmented white areas. Specifically, rats with two copies of the wild-type allele (HH) are solid colored, rats with one copy of each (heterozygous Hh) are solid with a white belly ("berkshire"), and rats with two hooded alleles have a solid colored head, a white body, and a colored dorsal stripe ("hooded").
Selection of rats for tameness results in depigmentation
Trut (1997) conducted the same experiment on rats as had been done on silver foxes. She used a group of wild agouti rats in captivity and selected them for tameness over generations.
Over 30 generations, the number of piebald (Hh) rats increased sharply in the population, until about 73% of the rats had white bellies. The white patches on the piebald rats grew larger and larger. The white pigment sometimes spread up the sides, and many of the animals had white feet. Over time, solid-colored rats disappeared from the population.
In a control population of rats not selected for tameness, a few piebald rats appeared as well, but they never comprised more than 10% of the population. The white spots of these piebald rats were small and confined to the chest or belly. They had no depigmentation on their sides and no white socks.
In sum, selection of rats for tameness correlated with the frequency and extent of depigmentation.
How the hooded gene regulates depigmentation
The hooded gene affects the distribution of pigment cells in the skin. Areas with pigment cells produce colored hairs, areas without pigment cells produce unpigmented, white hairs.
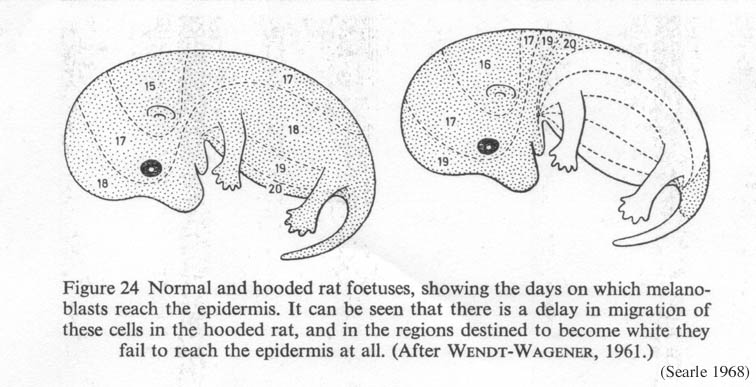
The story starts during development. In the embryo, a fold develops down the back called the neural crest. This region supplies cells that migrate all over the body and become a part of many different tissues. Anything that affects the migration of these cells is therefore going to have a wide variety of effects all over the body later in life.
Young pigment cells (melanoblasts) make this journey too. Pigment cells originate in the neural crest and normally migrate all over the body during development. Such animals produce pigment everywhere.
However, in rats with the hooded mutation, this migration of pigment cells is delayed (Wendt-Wagener 1961). Consequently, the areas of the body furthest from the back such as the feet, chest and belly don't have pigment cells, so those areas produce depigmented, white hair.
|
|
Normal and hooded rat fetuses, showing the days on which pigment cells reach different parts of the body (Searle 1968, from Wendt-Wagener 1961) Click on the image to get a larger view. |
How the hooded gene may influence behavior
Pigment cells aren't just found in the skin and hair follicles. They are also found in the central nervous system, including areas of the brain related to mood and response to stress.
Therefore, by selecting for docility in rats, humans select for changes in the biological systems that underlie behavior -- the hormonal and neurochemical systems. Changes in these systems have far-reaching effects.
In the case of the hooded mutation, selection for docility led to a change in developmental timing: a delay in migration of pigment cells. This delay changed the neural profile of the brain in areas related to mood and stress, and, incidentally, this delay caused a piebald coat.
Pigment cells also migrate to the inner ear and eye, and related neural crest cells migrate to the intestines. This is why there is often a relationship between blue eye color, odd-eyes, deafness, white coat or white blazes, and megacolon.
More on the mechanisms of coat color mutations.